Um alle Funktionen dieser Seite zu nutzen, aktivieren Sie bitte die Cookies in Ihrem Browser.
Meine Merkliste
my.chemie.de
my.chemie.de
Mit einem my.chemie.de-Account haben Sie immer alles im Überblick - und können sich Ihre eigene Website und Ihren individuellen Newsletter konfigurieren.
- Meine Merkliste
- Meine gespeicherte Suche
- Meine gespeicherten Themen
- Meine Newsletter
Michaelis-Menten-TheorieDie Michaelis-Menten-Theorie (nach Leonor Michaelis und Maud Menten 1913) legt den Grundstein für die Enzymkinetik. Hier wurde das theoretische Rüstzeug erarbeitet, Enzyme nicht nur hinsichtlich ihrer Aktivität zu charakterisieren, sondern auch die Stoffmenge (Konzentration) zu finden, welche eine den Gegebenheiten angepasste Umwandlung ermöglicht (siehe auch: Affinität (Biochemie)). Produkt-HighlightIm Allgemeinen sind Enzyme in der Lage, schwankende Substrat-Konzentrationen auszugleichen, d. h. ein Fließgleichgewicht („steady state“) dadurch einzustellen, dass sie ihre Tätigkeit dem Angebot anpassen. Ausnahmen hiervon bestätigen die Regel, und dann aus naheliegenden Gründen. So gibt es zwei Glucose-umwandelnde Enzyme, die Glucokinase der Leber und der Bauchspeicheldrüse und die Hexokinase, die in nahezu jeder Zelle vorkommt. Erstere trägt dazu bei, schädliche Schwankungen des Blutzuckerspiegels auszugleichen, indem 1. die Leber die Glukose aus dem Blut aufnimmt und in Form von Glykogen speichert und 2. je nach Glukosekonzentration im Blut eine angemessene Menge Insulin aus der Bauchspeicheldrüse sezerniert wird. Die Hexokinase hingegen arbeitet stets an ihrer Leistungsgrenze, das heißt auch bei niedrigem Blutzucker. Dadurch werden, besonders unter Mangelbedingungen, vorrangig das Hirn und die roten Blutzellen, die auf Glukose als Energiequelle angewiesen sind, mit Energie versorgt. Anders gesagt: die Hexokinase findet auch in Notsituationen stets noch Glucose, arbeitet also mit maximaler Effizienz. Sättigung: das PhänomenIm Gegensatz zur Kinetik chemischer Reaktionen gibt es in der Enzymkinetik das Phänomen der Sättigung: bei sehr hohen Substratkonzentrationen kann die Umsatzgeschwindigkeit v nicht weiter gesteigert werden d. h. es wird ein Wert Vmax erreicht. theoretische BegründungAls Biokatalysatoren bilden Enzyme E mit ihrem Substrat S einen Komplex ES, aus dem heraus sich die Reaktion zum Produkt vollzieht:
Hierin ist k2 ein Maß der maximalen Reaktionsgeschwindigkeit bei Substratsättigung ( Vmax), auch Wechselzahl, molekulare Aktivität, „turnover number“ oder kcat genannt (kcat = Vmax/ [Eo]). Die Michaeliskonstante, die Substratkonzentration, die bei der Halbsättigung vorliegt (die Umsatzgeschwindigkeit also v = Vmax/2 beträgt), ergibt sich zu (Michaelis-Menten-Fall, gegeben wenn k2<< k1) oder allgemeiner zu (Briggs-Haldane-Situation [Zerfallswege von ES/Bildungsweg von ES] für den Fall, dass k 2 gegenüber k 1 nicht vernachlässigt werden kann) Sättigungsverlauf: eine Hyperbel zwischen Km und Vmax
Die Sättigungsfunktion eines “Michaelis-Menten Enzyms” lässt sich unter Verwendung der Parameter Km und Vmax wie folgt formulieren: Dies ist die Michaelis-Menten Beziehung, d. h. die Gleichung einer Hyperbel mit den folgenden in der Abbildung gezeigten Eigenschaften:
Unter Enzymkinetik finden sich weitere Merkmale dieser Inhibitionstypen. |
| Dieser Artikel basiert auf dem Artikel Michaelis-Menten-Theorie aus der freien Enzyklopädie Wikipedia und steht unter der GNU-Lizenz für freie Dokumentation. In der Wikipedia ist eine Liste der Autoren verfügbar. |



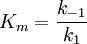
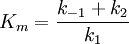
![v= {v_{max} \times [S] \over K_m + [S]}](images/math/1/1/d/11dcd4a06e793ee15318b1320b08a117.png)


