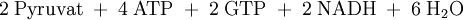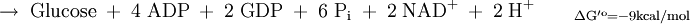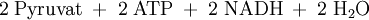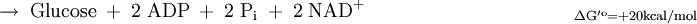Um alle Funktionen dieser Seite zu nutzen, aktivieren Sie bitte die Cookies in Ihrem Browser.
Meine Merkliste
my.chemie.de
my.chemie.de
Mit einem my.chemie.de-Account haben Sie immer alles im Überblick - und können sich Ihre eigene Website und Ihren individuellen Newsletter konfigurieren.
- Meine Merkliste
- Meine gespeicherte Suche
- Meine gespeicherten Themen
- Meine Newsletter
GluconeogeneseDie Gluconeogenese (latinisierte Schreibung der Glukoneogenese, eines Kompositums aus altgriech. γλυκύς, „süß“, νέος, „neu“ und γέννησις, „Erzeugung“) ist eine Neusynthese von Glucose aus organischen Nicht-Kohlenhydrat-Vorstufen wie Pyruvat, Oxalacetat und Dihydroxyacetonphosphat. Dieser Stoffwechselweg ist ubiquitär verbreitet, jedoch existiert in Pflanzen und Bakterien eine weitere Möglichkeit aus organischen Vorstufen (in dem Fall Acetyl-CoA) Glucose herzustellen, der Glyoxylatzyklus. Produkt-Highlight
Notwendigkeit der Gluconeogenese beim MenschenDer tägliche Glucosebedarf eines Erwachsenen Menschen beträgt ungefähr 160 g, wobei davon allein 120 g vom Gehirn genutzt werden. Die verfügbare Menge an Glucose im Blut entspricht circa 20 g (Plasmaglukose), die Menge an Glycogen, die im Körper gespeichert ist, beträgt etwa 400–450 g. Davon sind ca. 350 g in der Muskulatur gespeichert und ca. 80–100 g in der Leber. Die Erythrozyten sind, da die einzigen menschlichen Zellen ohne Mitochondrien, auf die Zufuhr von Glucose angewiesen. Das Gehirn dagegen wegen seines enormen Bedarfs an schnell verfügbarer Energie. Deshalb v.a. setzt bereits bei relativ kurzfristigen Hungerperioden die Synthese von Glucose ein. Diese findet vor allem in der Leber und in der Nierenrinde statt, weniger im Gehirn, Skelett- und Herzmuskel. Pro Tag können etwa 180–200 g Glucose gebildet werden. Ablauf der GluconeogeneseDie Ausgangsstoffe der Gluconeogenese werden entweder in Form von Pyruvat oder Oxalacetat, als Produkte des Aminosäureabbaus und der Milchsäuregärung (aus Lactat), oder in Form von Dihydroxyacetonphosphat, als Derivat von Glycerin aus dem Fettabbau, in den Stoffwechselweg eingeschleust. Die Gluconeogenese entspricht nur teilweise der Umkehrreaktion der Glykolyse, in der das chemische Gleichgewicht dreier Teilreaktionen fast ausschließlich auf der Seite der Reaktionsprodukte liegt. Die anderen Umwandlungsprozesse befinden sich im Gleichgewicht, weshalb diese auch bei der Gluconeogenese eine Rolle spielen. Die für die Glykolyse typischen Reaktionen sind:
Diese Schritte werden in der Gluconeogenese in umgekehrter Reihenfolge wie folgt umgangen:
Ein weiterer wichtiger Unterschied zur Glykolyse ist der Reaktionsort. Während diese ausschließlich im Cytosol abläuft, ist die Gluconeogenese auf drei Kompartimente verteilt. Die Umwandlung von Pyruvat in Oxalacetat erfolgt im Lumen des Mitochondriums, weshalb das Malat Shuttle-System für den Transport des Oxalacetats aus dem Mitochondrium ins Cytosol verantwortlich ist. Auch der letzte Reaktionsschritt findet nicht im Cytosol statt, sondern im Endoplasmatischen Retikulum (ER). Hierbei spielt die Glucose-6-phosphatase eine Rolle, ein membrangebundener Enzymkomplex aus mehreren Einheiten, die auch Kanäle für die an der Reaktion beteiligten Stoffe bilden. Pyruvat-Carboxylase
Die erste Teilreaktion ist abhängig von der Anwesenheit von Acetyl-CoA, d.h. ohne dieses ist keine Carboxylierung von Biotin möglich. Diese Regulation ist eine Form von Allosterie, da ein hoher Acetyl-CoA-Spiegel ein Zeichen für mehr Bedarf an Oxalacetat im Citratzyklus steht. Bei ATP-Überschuss jedoch wird das Oxalacetat in der Gluconeogenese verbraucht, wodurch dieses nicht angereichert wird. Diese Funktion der Pyruvat-Carboxylase nennt man anaplerotisch, d.h. diese Enzym liefert wichtige Metabolite nach. Der zweite Reaktionsschritt der Pyruvat-Carboxylase ist Acetyl-CoA unabhängig. Vergleich Gluconeogenese und GlykolyseEnergiebilanz im Vergleich zur Umkehrung der GlykolyseFür die Biosynthese von einem Molekül Glucose werden ausgehend vom Pyruvat vier Moleküle ATP und je zwei Moleküle GTP und NADH benötigt.
Durch die unten aufgeführte Bilanz wird deutlich, dass die obere Reaktion bevorzugt ablaufen wird, da eine direkte Umkehrung der Glykolyse eine thermodynamisch ungünstige Reaktion darstellt.
Gluconeogenese und Glykolyse – reziproke RegulationDie Gluconeogenese wird an zwei Stellen reguliert:
Zur ersten Reaktion: die in der Glykolyse vorkommende Umwandlung von PEP in Pyruvat wird von der Pyruvatkinase katalysiert. Die Aktivität dieses Enzyms wird durch Fructose-1,6-bisphosphat erhöht und durch ATP und Alanin inhibiert. Die Enzyme der Gluconeogenese (Pyruvatcarboxylase und PEP-carboxykinase) werden durch Acetyl-CoA aktiviert und durch ADP gehemmt. Da ATP durch Hydrolyse in ADP umgewandelt wird, kann man bei dieser Art der Regulation zweier gegenläufiger Reaktionen von reziproker Regulation sprechen. Ein weiteres Beispiel bietet hierfür die unter 2. aufgeführt Reaktion. Die bei der Glykolyse beteiligte Phosphofructokinase wird durch Fructose-2,6-bisphosphat und Adenosinmonophosphat (AMP) stimuliert, jedoch unter anderen durch Citrat inhibiert. Reziprok dazu findet die Regulation der an der Gluconeogenese beteiligte Fructose-1,6-bisphosphatase statt (durch Citrat aktiviert und durch Fructose-2,6-bisphosphat und AMP gehemmt). Siehe auchGlyoxylat-Zyklus, Fructose-6-phosphat, Cori-Zyklus Schemata
Kategorien: Biochemische Reaktion | Stoffwechsel |
|||
| Dieser Artikel basiert auf dem Artikel Gluconeogenese aus der freien Enzyklopädie Wikipedia und steht unter der GNU-Lizenz für freie Dokumentation. In der Wikipedia ist eine Liste der Autoren verfügbar. |



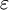 -Aminogruppe eines spezifischen Lysinrestes gebunden. Dadurch entsteht ein flexibler Arm, wodurch die Biotingruppe von einem aktiven Zentrum zum zweiten "schwingen" kann. Die Carboxylierung erfolgt in zwei Schritten:
-Aminogruppe eines spezifischen Lysinrestes gebunden. Dadurch entsteht ein flexibler Arm, wodurch die Biotingruppe von einem aktiven Zentrum zum zweiten "schwingen" kann. Die Carboxylierung erfolgt in zwei Schritten:
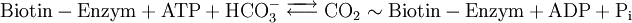
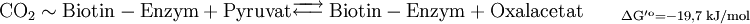 .
.